My research interests lie in mathematical modeling and analysis of biological phenomena. My research efforts to date can be divided into two major components -- multi-scale modeling of colon tissue cell dynamics and colorectal cancer initiation, and semi-discrete modeling of host-parasitoid population dynamics. My professional goal is to continue to apply mathematical techniques to biological problems, with an emphasis on collaborative research and undergraduate mentorship. Below are several figures that summarize some past and present research projects.
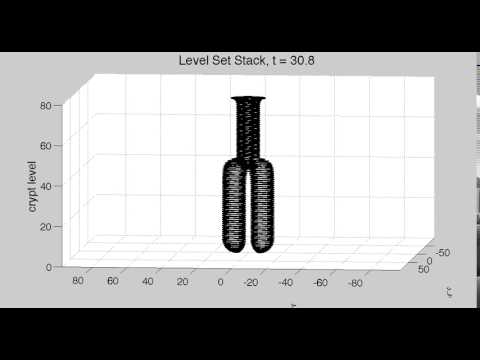
Cell Dynamics of the Colonic Crypt
Hybrid Systems of Host-Parasitoid Ecology




Other Interesting Projects


The Wnt Pathway is a cascade of chemical reactions within a cell that ultimately determines the destruction or accumulation of a proliferation promoting protein called beta-catenin. This figure shows a single trajectory in the phase space of a system of nonlinear differential equations that describes the interactions of three key proteins involved the Wnt pathway: beta-catenin, Axin, and APC. When the concentration of Axin is relatively small, the three concentrations become periodic in time. This indicates that a cell switches, periodically, between a proliferative and quiescent state. The paper is here.